 Look at the head and jaws of a halfbeak:
Look at the head and jaws of a halfbeak:
The lower jaw appears very elongate, reaching as much as ten times further than the upper jaw. What is the function of this bizarre anatomy? Is it used in feeding? Is it used in locomotion? If so, how?
Halfbeaks (Hemiramphidae) are related to needlefish, flying fish and sauries. They are slender-bodied fish that spend most of their time swimming right up at the surface of the water where they are expert at hiding from predators. They mainly occupy warm and temperate inshore marine habitats, though there is also a radiation of freshwater forms. The marine species feed on drifting pieces of plants and zooplankton. The freshwater species and most inshore marine forms also eat drifting surface insects.
Once you get past the very strange appearance of these fish and closely inspect the head you find that the long lower jaw is not actually a lower jaw but instead an absurdly long chin (have a look at the diagrams below from Montgomery & Saunders 1985). The toothed lower jaw is short and matches the toothed upper jaw. The halfbeak is actually a very long extension of the part of the mandible that is below, and in front of the teeth – a chin. The structure is smooth and not armed with teeth. This is our first clue that the structure is not an elaborate jaw used in prey capture. If it’s not part of the feeding apparatus, then how does it function?
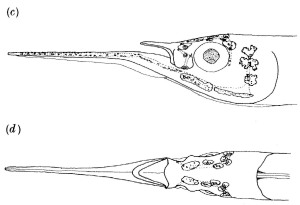
One idea that has some support is that the long chin is part of a specialized sensory devise. Montgomery & Saunders (1985a) showed that there are a series of lateral line pores along the length of the lower jaw, with neuromasts in between these pores. They argued that this long structure, equipped with the lateral line pits, may function in prey detection. The species they worked with feeds at night on large zooplankton and they showed it is capable of detecting live plankton in total darkness. In effect, they argue that the halfbeak functions as an extension of the lateral line system that may aid these fish in nocturnal attempts to locate moving prey animals in midwater habitats.
This is a fascinating proposal that may help us understand the function and evolution of the halfbeak. But the system requires more attention before we fully understand how it works and how it evolved. It would be valuable to model the potential sensory advantage of the long, sensitive chin. Does it convey a major performance advantage over fish that lack such a device? What was the evolutionary sequence here? Did the long chin evolve as a specialization of the lateral line system, or did the long chin evolve first for a different function, and then become co-opted for use as a scaffolding for the sensory system? And, what role does the long lower jaw play during prey capture? Such a large structure must interact with prey during feeding. Are there other, secondary functions of the long chin? What do you think?
Montgomery, J.C. & A.J. Saunders. 1985. Functional morphology of the piper Hyporhamphus ihi with reference to the role of the lateral line in feeding. Proc. Roy. Soc. Ser. B. 224:197.
Saunders, A.J. & J.C. Montgomery. 1985. Field and laboratory studies of the feeding behavior of the piper Hyporhamphus ihi with reference to the role of the lateral line in feeding. Proc. Roy. Soc. Ser. B. 224:209.


Recent Comments